
Índice de temas anteriores
Introducción
En esta tercera edición vamos a cerrar el círculo de los conceptos básicos y establecer las bases del circuito nervioso más sencillo: un reflejo medular de tan solo dos neuronas. En definitiva, hoy nos centraremos en:
- Cómo se forman los nervios periféricos
- Histología de un nervio modelo
- Concepto de dermómero
- Placa motora
- Receptores musculares
- Tipos de fibra nerviosa
- Reflejos medulares I: reflejo miotático
Por supuesto, de acuerdo con la resolución que tomé en el tema anterior, voy a reducir la carga de de contenidos de cada tema en aras de hacerlos más manejables y atractivos. Vamos allá.
Formación de nervios periféricos desde nervios espinales
Llevo dos lecciones eludiendo aclarar la diferencia entre nervio y fibra porque no es tan sencillo como asumir que una fibra = nervio. Para comenzar, recordad que para cada nivel medular existe un par de nervios espinales, por lo tanto, 31 pares. Los nervios craneales son especiales y vamos a obviarlos por ahora.
Los nervios espinales son el origen del inmenso árbol que abarca todo nuestro cuerpo, a excepción de la cabeza. Están formados por miles de axones de un nivel medular correspondiente. Primero, vamos a entender cómo se ramifican, se unen y se separan. Luego, entraremos a ver la composición de un nervio individualmente.
El origen de los nervios espinales: Raíces medulares

Como ya sabréis, esto es un corte axial de la columna vertebral en el que se aprecia la vértebra, el canal raquídeo y la médula espinal, por supuesto. Si os fijáis en el nervio raquídeo o espinal, observaréis que de la médula parten un par de raicillas que se fusionan en una única estructura, el nervio raquídeo. Llamaremos a estas raicillas raices espinales. A la que se encuentra más "abajo" en la imagen o ventral la llamaremos raíz anterior y a la que se encuentra justo más arriba o dorsal la llamaremos raíz posterior o dorsal.
Además, quiero que os fijéis que la raíz dorsal se ensancha formando una bolita o ganglio de la raíz dorsal (DRG para los amigos) del que parte otro haz que ya se une a la raíz anterior. Pues bien, os adelanto que en este ganglio se encuentra el soma de las fibras sensitivas de todo el nivel medular. Estas neuronas son pseudomonopolares, con una prolongación que se dirige a la periferia y otra, a la médula espinal. Os pido encarecidamente que repaséis el término pseudomonopolar en el tema anterior. Si la raíz posterior transporta fibras sensitivas, la raíz anterior lleva fibras motoras.
En resumen, todo nervio espinal tiene por origen un par de raíces espinales, una anterior y otra posterior. La raíz posterior forma primero un ganglio llamado ganglio de la raíz dorsal. La raíz dorsal o posterior transporta fibras sensitivas. La raíz anterior, motoras. El nervio espinal es ya, mixto.
El nervio torácico, el nervio más sencillo
Antes de meternos con los plexos, quiero explicaros el prototipo de nervio más sencillo. Uno en el que un nervio espinal simplemente abandona la columna, se divide en un ramo dorsal y otro ventral, ambos mixtos, con fibras sensitivas y motoras. El ramo posterior se dirige a la espalda y el ramo anterior se prolonga, ramificándose poco a poco, a lo largo de un segmento corporal muy bien definido. Por motivos que no vienen al caso, digamos que el tórax mantiene cierta "segmentación primigenia".
Una imagen para ilustrarlo:
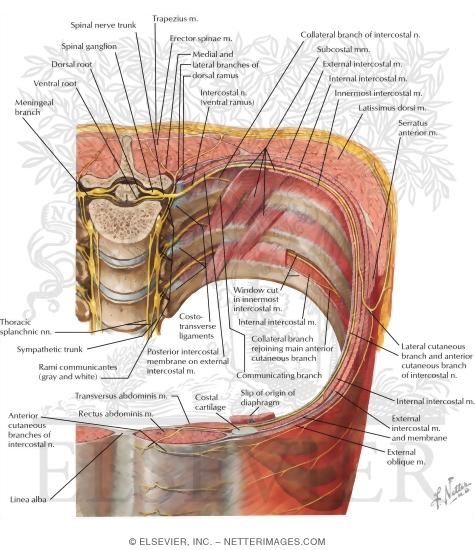
Tal y como se muestra en la imagen, de cada nivel medular parten un par de nervios torácicos que siguen un patrón segmentario repetitivo, ramificándose a lo largo del segmento que están bastante bien definidos. Más cuando hablemos de dermómero.
Plexos, las grandes autopistas
Como podéis imaginar, si bien el tórax conserva la citada segmentación original, no ocurre lo mismo con, por ejemplo, los hombros o los brazos. No voy a explicaros la biología del desarrollo que sirve como base a este hecho, así que tomadlo como parte de la anatomía humana: muchos nervios espinales se mezclan con otros nervios espinales, formando a su vez otros nervios que transportan fibras de varios niveles medulares.
Comencemos definiendo claramente un plexo. Un plexo es una agrupación de axones provenientes del ramo anterior de varios nervios espinales. Tenemos varios plexos, destacando por ejemplo el plexo braquial o el lumbar.
Otra imagen, esta del plexo cervical para que os hagáis una idea:
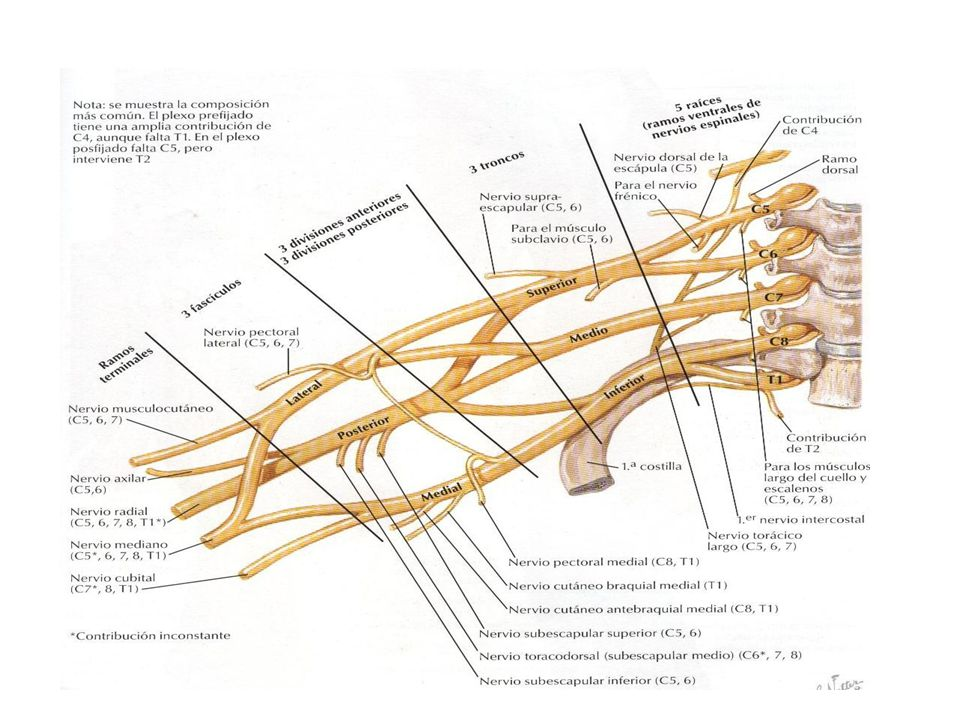
Este plexo origina todos los nervios que controlan el brazo. Le llamamos plexo braquial. Como veis, al final del plexo se forman varios nervios, como el mediano, que son el resultado de la unión de varios nervios espinales provenientes de varios niveles medulares.
Como esto no es un curso de Anatomía regional, no os voy a machacar más con esto.
Histología de un nervio modelo
Imaginad que cortamos un nervio cualquiera y lo miramos al microscopio. ¿Qué narices hay ahí? De eso vamos a enterarnos en esa sección.
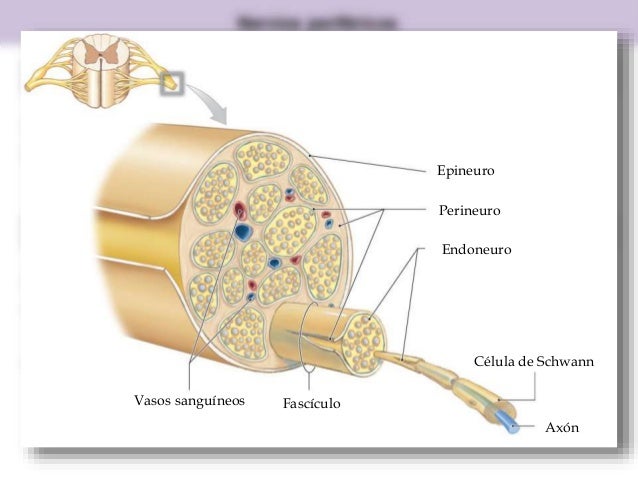
Este amable esquema nos servirá como base y sobre ella articularemos toda la sección. Terminaremos viendo algunas imágenes reales, como no. Por lo pronto quiero que os fijéis que todo nervio está rodeado por una vaina protectora, conjuntiva, llamada epineuro. El interior del nervio está formado por una serie de cúmulos nerviosos inmersos en una matriz conjuntiva y vasos sanguíneos. A cada cúmulo lo llamaremos haz o fascículo nervioso. Cada haz nervioso está delimitado y recubierto por otra vaina conjuntiva, el perineuro.
Finalmente, cada haz nervioso está formado por un conjunto de fibras o axones nerviosos, rodeados individualmente por ooooootra vaina llamada endoneuro. Recapitulando, un nervio es una estructura rodeada por epineuro, que a su vez está formado por varios haces nerviosos rodeados por el perineuro, que a su vez están formados por miles de fibras individualizadas y rodeadas por el endoneuro.
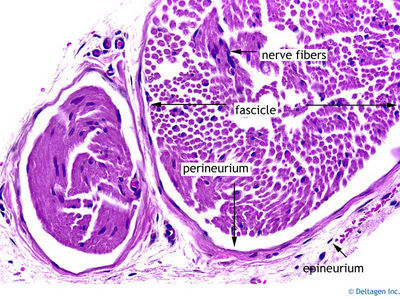
Y ahora unas imágenes histológicas para que veáis que no me invento las cosas:

Concepto de dermómero
Los dermómeros son muy sencillos. Si todo nervio tiene como origen un nervio espinal de un nivel medular correspondiente, es de traca que cada punto de nuestra piel está inervado por un nivel medular concreto. A la porción de la piel que se corresponde con un nivel medular concreto lo llamamos dermómero.
¡Vamos a verlo!

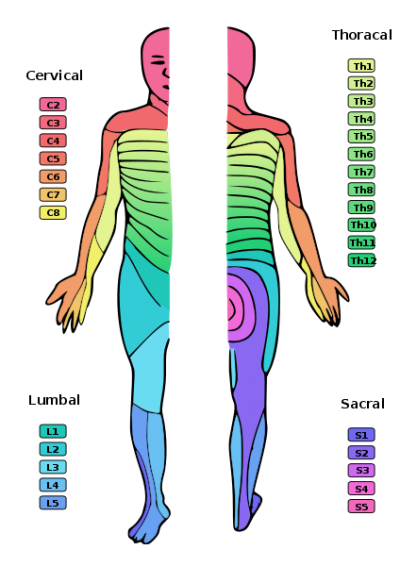
Fijáos que el tórax, que mantiene muy bien la "segmentación primigenia", tiene unos dermómeros que prácticamente son aros que rodean al nivel medular correspondiente. Sin embaaaaaaargo, allá donde hay un plexo, como el braquial, nos encontramos con dermómeros más irregulares. Las extremidades son el lugar donde más se distorsiona la segmentación, pero bueno, ya os dije que no hablaríamos de biología del desarrollo.
Recordad que, dermómero se refiere a nivel cutáneo SOLA Y EXCLUSIVAMENTE. Además, esta división es aproximada y los bordes entre cada dermómero son difusos. Más cuando hablemos de campos.
Placa motora, la base del movimiento
Hasta ahora hemos estudiado como los axones de las neuronas viajan por el cuerpo, pero ignoramos aún de dónde vienen los estímulos o a dónde van. En esta sección voy a explicaros cómo se pasa de un potencial de acción a una contracción muscular. Si recordáis el principio del tema, os hablé de que la raíz anterior aportaba fibras motoras o motoneuronas, esto es, axones que son capaces de producir la contracción de la musculatura. ¿Cómo lo hacen?
Todo comienza con un potencial de acción en una motoneurona, a la que llamaremos segunda motoneurona -conoceréis a la primera más adelante-, cuyo axón o fibra motora viaja por la raíz anterior del nervio espinal correspondiente hasta el músculo que toca. Efectivamente, a cada músculo le corresponden uno o más niveles medulares.
Este potencial de acción llega hasta una unidad motora, la unidad mas básica de la contracción.
Unidad motora
Una unidad motora no es más que el conjunto formado por una fibra motora y las fibras musculares que inerva. Lamentablemente, no es objeto de este curso entrar en la histología de la musculatura, por lo que os tendréis que conformar con imaginar cada fibra como una cuerda capaz de contraerse y al músculo como un conjunto de cuerdas. Cuando una fibra se excita, produce la contracción simultánea de todas las fibras que inerva, trabajando juntas.
Un par de esquemas para que lo entendáis


La placa motora: la unión neuromuscular
DISCLAIMER: LA CONTRACCIÓN MUSCULAR DARÍA PARA UN TEMA ENTERO EN SÍ MISMO. Incluyendo la histología y ultraestructura de la célula muscular. Yo os presento una simplificación muy simplificada. Narices, la fisiología del calcio en la célula muscular da para vaaaarios temas.
Una placa motora, como podréis imaginar, es el análogo a una sinapsis entre una fibra motora y una fibra muscular. Cuando el potencial de acción alcanza la terminal axónica, se produce la liberación de neurotransmisores tal y como ya os expliqué en el tema anterior. Sin embargo, estos neurotransmisores no producen otro potencial de acción, sino una onda de calcio que hace posible un evento contráctil.
El neurotransmisor que libera la terminal axónica es acetilcolina, el cual se une a los receptores de la membrana de la fibra muscular. Estos receptores de acetilcolina son nicotínicos. Afortunadamente para vosotros, su funcionamiento es sencillísimo. Estos receptores son canales de Na dependientes de ligando -tachán, un receptor puede ser un canal- por lo que entra Na en la membrana de la fibra muscular.
La entrada de Na en la célula muscular la despolariza, lo cual hace que se abran unos canales de calcio de tipo L y que además son voltaje dependientes, como podréis imaginar. Le vamos a dar un nombre especial a estos canales: dihidropiridínicos o de dihidropiridina. Bien, la entrada de calcio desde el exterior de la célula muscular es INSUFICIENTE para causar la contracción.
Sin embargo el canal de dihidropiridina está acoplado a otro canal de calcio, al cual llamaremos de rianodina. Están tan acoplados que son una única unidad, de forma que cuando se abre uno se "fuerza" la apertura del segundo. Estos últimos canales están a su vez conectados a un gran depósito de calcio llamado retículo sarcoplásmico que es, como no, un gran depósito de calcio.
La onda de calcio es la que señala que la contracción ha de comenzar.
Un esquema para aclararlo todo:

En resumen, el potencial de acción produce la liberación de acetilcolina, que a su vez actúa como molécula señal para la contracción muscular. Todo comienza con la entrada de Na, que produce una despolarización que a su vez permite la liberación de Ca del gran depósito llamado retículo sarcoplásmico.
Si queréis profundizar y que nos metamos a hablar desde el principio de la contracción muscular -sería un tema entero- comentadlo y lo miramos. Os advierto que es sencillo pero largo.
Reclutamiento fibrilar
Sólo comentaros un par de conceptos acerca de este fenómeno fisiológico. Si un músculo está formado por unidades motoras y un nervio puede contener cientos de axones, no es de extrañar que a una unidad motora puedan sumarse otras, estimuladas por otros axones, aumentando la fuerza de la contracción. Esto es, en pocas palabras, lo que significa reclutar unidades motoras. Por lo tanto, cuantas más motoneuronas se activen, más unidades motoras se contraen y más fuerza se ejerce. ¿Cómo se coordinan unas y otras? Poco a poco, poco a poco... Por ahora me conformo con que sepáis que existe.
Receptores musculares
Ya sabemos cómo se produce la contracción muscular, la base del movimiento. Ahora vamos a centrarnos en cómo nuestro sistema nervioso percibe la contracción muscular y por tanto, el movimiento . Mi intención inicial era incluir en este tema receptores superficiales y campos sensitivos, pero pensándolo mejor voy a dejarlo estar por ahora. Habrá tiempo de sobra cuando hablemos de reflejo de retirada, termoalgesia...
Por tanto, vamos a estudiar los husos musculares y el órgano de Golgi.
Huso muscular
A este receptor se le llama huso muscular porque tiene forma de huso y además, se encuentra en el interior de los músculos.
En efecto, el huso muscular es un receptor profundo que detecta cambios en la longitud del mismo. En otras palabras, cuando el músculo se alarga, se activa. Gracias a ellos podemos detectar además la fuerza de la contracción, en función de la variación en la longitud del músculo y regularla. Más adelante cuando hablemos del reflejo miotático lo veréis.
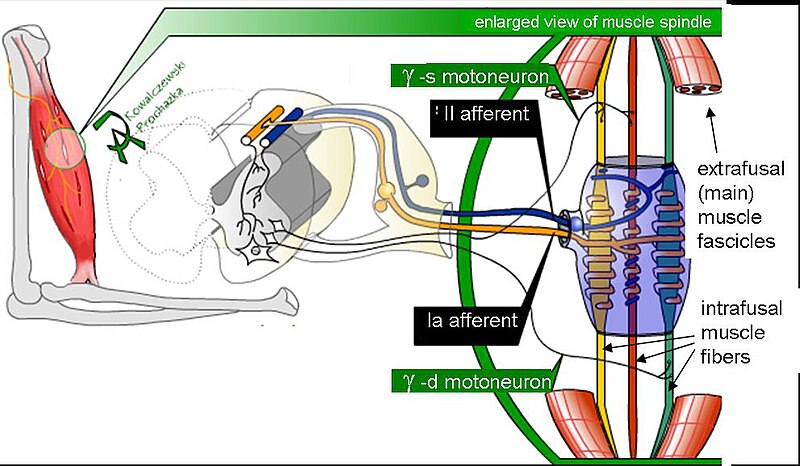
NO OS ASUSTÉIS. El esquema es más fácil de lo que parece. Fijaos que han cortado dos fibras musculares, entre las cuales se encuentran otras fibras más pequeñas o intrafusales. Alrededor de ellas se encuentran las terminales axónicas de dos neuronas sensitivas pseudomonopolares. Una amarilla y otra azul. Ambas tienen su soma -la bolita- en el ganglio de la raíz dorsal... ¡cómo todas las neuronas sensitivas! y se dirigen a la médula espinal.
¿Cómo funciona este receptor?
Cuando el músculo se estira, se activan canales de Na en la terminación nerviosa de manera mecánica, por estiramiento del huso.
Fijaos además que la fibra amarilla rodea la porción más gruesa de la fibra intrafusal y la fibra azul sólo los extremos. Además hay una tercera fibra intrafusal que no tiene un vientre regordete. El significado funcional de unas y otras no es objeto de este curso. Una se llama Ia y otra II, me conformo con que recordéis que un huso muscular está inervado, bien por una fibra Ia, bien por una II
Sí me interesa que os pispéis del hecho de que hay una fibra motora eferente que está inervado estas fibras intrafusales. En efecto, es una motoneurona y en efecto, estas fibras se contraen. Pero el objetivo de estas fibras delgaduchas no es aumentar la fuerza de la contracción, sino regular la sensibilidad del receptor. A las fibras motoras a las que nos estamos refiriendo las llamaremos gamma o fusimotoras.
¿Cómo lo hacen?
¿Os acordáis de que para que se inicie un potencial de acción hace falta que entre suficiente Na? Pues muy sencillo. La contracción, gracias a las fibras gamma o fusimotoras, de la fibra intrafusal hace que los extremos se acorten. Sin embargo, la parte gordita no puede contraerse. ¿Resultado? Es como si dos hermanos tiran de un juguete, cada uno hacia un extremo: la parte gordita se alarga.
Alrededor de esta parte gordita están las fibras aferentes, enrolladas. La elongación activa canales de Na, lo cual hace que suba el potencial de reposo de la fibra, pero ojo, no lo suficiente como para que se desencadene un potencial de acción. Lo que sí pasa es que facilita que un estímulo contráctil menos intenso por parte de las fibras extrafusales produzca una despolarización. Ergo, cuando las fibras intrafusales se contraen, el receptor se hace más sensible y viceversa.
Con estiramientos más suaves, tendríamos potencial.
Órgano tendinoso de Golgi
NO LO CONFUNDÁIS con el aparato de Golgi. Esto es un receptor. En concreto, un receptor profundo que se encarga de detectar el estiramiento de los tendones, en el origen y el final de cada músculo. Es por tanto muy parecido al huso muscular. De hecho, trabajan juntos.
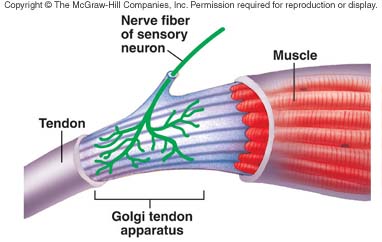
Por suerte para vosotros también son bastaaaante más sencillos. Reservaros las bromas entre GTO (Golgi tendon apparatus) y GTFO y reunid la poca concentración que os quede para el último de los receptores de hoy. Imaginaos que entre todo el colágeno que forma un tendón hay una serie de fibras que, saliendo del final del músculo, forman "aros" en los que se inserta la fibra nerviosa.
Cuando el músculo se contrae, el tendón se alarga, lo cual reduce la horquilla de la fibra de colágeno, estrangulando la terminal nerviosa y activando canales de Na que producen un potencial de acción. Un esquema sencillo os lo aclarará todo:

Al igual que me interesaba que recordarais las fibras Ia y II, ahora quiero que recordéis que cada órgano de Golgi -hay muchos por tendón- está inervado por una única fibra, que llamaremos Ib.
Tipos de fibra nerviosa
Esta tabla ya la habéis visto, pero ahora estamos en disposición de entender UNO de los grupos. No os preocupéis por el resto, los vamos a volver a ver. Es más, os acabaréis aprendiendo la clasificación de memoria a base de pura repetición. Crearé el apartado de "tipos de fibra" tantas veces como sea necesario y así poco a poco iréis construyendo vosotros mismos la tabla que ahora os muestro:

¿Qué fibras hemos visto hoy?
La segunda motoneurona del músculo esquelético -lo que vosotros entendéis por músculo vaya- es una fibra tipo A, subtipo alfa. Como veis son del tipo más grande y más mielinizado, por tanto más rápido. Hasta 80 m/s. Los husos musculares son fibras Ia y Ib, es decir, una parte de las fibras aferentes que vienen del huso muscular y todas las del órgano de Golgi son también de tipo A, subtipo alfa.
Las fibras tipo II, que también vienen del huso muscular, son sin embargo un poco más pequeñas y menos mielinizadas, por lo tanto más lentas. Pertenecen al grupo A, subgrupo Beta. Por último, las fibras eferentes gamma o fusimotoras forman ellas solitas el grupo A, subgrupo gamma.
Reflejos medulares I: reflejo miotático
Y después de tanta cháchara vamos por fin a ver NUESTRO PRIMER CIRCUITO NERVIOSO. El reflejo miotático es el circuito más simple de todos, ya que solo necesita de dos neuronas para funcionar y se encuentra enteramente en la médula espinal. ¡Funcionaría incluso si os cortase la cabeza!
Antes de nada y aunque en el siguiente tema volveré a hacerlo, os adelanto ya que un reflejo es una respuesta automática e involuntaria que realiza un ser vivo ante un determinado estímulo. Normalmente consiste en un movimiento, aunque también hay reflejos secretores. Algunos autores consideran los reflejos medulares como el tipo más primitivo de memoria.
¿Qué son los reflejos medulares? Aquellos en los que los arcos o circuitos se encuentran en la médula espinal. Son, por lo general, movimientos extremadamente rápidos. Consideradlos como respuestas que deben iniciarse tan rápidamente que ni siquiera se espera al procesamiento del estímulo. Conforme llega, comienza la respuesta.
El reflejo miotático
Este reflejo, el más simple de todos, también se conoce como reflejo de estiramiento. Es un regulador automático del estiramiento muscular. Cuando un músculo se estira más allá de cierto punto, este reflejo contrae automáticamente el músculo para evitar una rotura. Os lo habrán explorado mil veces con un martillo en la rodilla.
¿Cómo ocurre?
El receptor de estiramiento muscular es, lógicamente, el huso muscular, ya que es el encargado de detectar cambios en la longitud, bien acortamiento, bien estiramiento, del músculo. En su forma más simple, el reflejo miotático consiste en la activación de las fibras Ia y II, que a su vez activan a una motoneurona en la médula para que contraiga una unidad muscular. Así, el músculo se acorta y se restablece la longitud.
Un esquema nunca hace daño:

Cuando nos dan con el martillito en la rodilla, ocurre lo mismo que si se activaran los músculos isquiotibiales. Al activarse estos músculos, la pierna se estira y el cuádriceps se estira a su vez. El martillo, al tirar del tendón del cuádriceps, hace exactamente lo mismo, estirarlo. Como veis esto activa a una neurona pseudomonopolar que inerva los husos del cuádriceps, que va a la médula y activa mediante sinapsis química a una motoneurona -la que llamamos segunda motoneurona- que controla una unidad muscular en el cuádriceps. El cuádriceps se contrae en respuesta, recuperando su longitud.
Este reflejo es monosináptico, esto es, sólo hay una sinapsis. Para cada fibra aferente, una motoneurona. Por eso es tan sumamente rápido. En sentido estricto, en esto consiste el reflejo miotático.
Peeero, observaréis además que la misma neurona aferente da una segunda rama a otra neurona. Esa segunda neurona, roja y de axón corto, es una interneurona de la médula espinal. En este caso, la activación de la interneurona hace una cosa muy curiosa, ya que produce una INHIBICIÓN de otra motoneurona. ¿Recordáis las sinapsis inhibidoras del tema anterior? Efectivamente, hacen más difícil que la motoneurona dispare. ¿Y a donde va esa motoneurona? A los isquiotibiales, los antagonistas del cuádriceps.
Por lo tanto, el reflejo miotático ampliado aplicado al cuádriceps consiste en la activación del cuádriceps -que se contrae en respuesta al estiramiento- y la relajación del bíceps femoral, que al contraerse ¡estiraría el cuádriceps!
Y así, amigos míos, es como se regula -entre otros mecanismos- el tono muscular entre agonistas y antagonistas. Si un músculo se contrae con demasiada fuerza, el músculo antagonista -el que realiza el movimiento contrario- se estira, lo que activa el reflejo miotático, que tiende a inhibir la contracción de uno y a contraer el contrario.
Últimas palabras
Este tema es más corto y sencillo, por lo que no deberíais tener problemas. El siguiente tema lo dedicaré en exclusiva a reflejos medulares, así que id quedándoos con la nomenclatura de bolitas y líneas de circuitos. Repetiré y ampliaré el término reflejo. Como siempre si quedan dudas ya sabéis.
PD: A lo largo de mañana haré algunos ajustes en la redacción, si veis cualquier gazapo o argumento circular fruto de trasnochar avisadme y lo arreglo ASAP.

